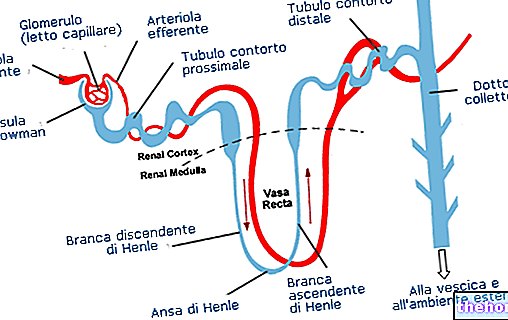
"перша частина
Ферментом, який каталізує четверту стадію циклу Кребса, є α-кето-глутаратдегідрогеназа; цей фермент є ферментним комплексом, дуже схожим на піруватдегідрогеназу. Обидва складаються з 48-60 білків, у яких розпізнаються три різні ферментативні дії, а також вони мають однакові ферментативні кофактори; дуже схожі ферменти, оскільки вони діють на подібні субстрати: і піруват, і л "α-кето глутарат, є α- кетокислоти. Механізм дії двох ферментативних комплексів однаковий.
Напад тіамінпірофосфату на карбоніл (С = О) "α-кетоглутарат, призводить до його декарбоксилювання та утворюється похідне карбоксигідроксипропілу. З подальшим переходом до ліпоаміду відбувається внутрішній окислювально-відновний процес, з якого отримують похідне ліпоаміду карбокси або сукцинілліпоамід.
Потім сукциніловий ліпоамід реагує з коензимом А з утворенням сукцинілового коферменту А (який триває у циклі Кребса) та відновленого ліпоаміду, який повторно окислюється FAD: утворюється FADH2 окислюється за допомогою НАД + та НАДН. Тому на цій стадії відбулося друге видалення вуглецю з карбонового скелета у вигляді вуглекислого газу.
Ацильна група, пов'язана з коферментом А, знаходиться в активованій формі, тобто вона має високий вміст енергії: отже, можна використовувати енергію сукцинілового коферменту А.

На п'ятій стадії циклу Кребса сукциніловий кофермент А піддається дії сукцинілтіокіназа; було висунуто дві гіпотези щодо її способу дії: ми опишемо лише одну з двох, оскільки вона є найбільш акредитованою. Згідно з цією гіпотезою, сукциніл-кофермент А піддається впливу азоту гістидину (Hys) ферменту: виділяється коензим А і утворюється адукт, отриманий з гістидину, як проміжний продукт, тобто сукциніл-фермент ); на цей проміжний продукт діє ортофосфат, що призводить до вивільнення сукцинату та утворення фосфоензиму. З точки зору енергії GTP = АТФ: зв’язок, що забезпечує енергію, однаковий у обох видів (це ангідридний зв’язок між фосфорилом Β та фосфорилом γ). У деяких випадках ГТФ використовується як матеріал з високим вмістом енергії, але зазвичай ГТФ перетворюється в АТФ під дією ферменту нуклеозиддифосфокіназа (НДПК); - фермент, що міститься в клітинах і каталізує наступну реакцію:
N1TP + N2DP → N1DP + N2TP
Загальний нуклеозидтрифосфат NiTP ®
Загальний нуклеозид дифосфат NiDP ®
Це оборотна реакція; в нашому випадку це відбувається:
GTP + ADP → GDP + ATP
тому він може рухатися вправо або вліво навіть при невеликих коливаннях концентрацій реагентів.
Якщо цикл Кребса протікає з такою швидкістю, що призводить до вироблення АТФ, вищого за енергетичну потребу, існує недостатня кількість АДФ, а АТФ багато: реакція, каталізована нуклеозиддифосфокіназою, спрямовані вліво (GTP накопичується, якщо нуклеозидна дифосфокіназа не має достатнього субстрату, тобто АДФ). Тому GTP є сигналом доступності енергії і тому уповільнює цикл Кребса.
Шоста стадія циклу Кребса призводить до утворення фумарату під дією сукцинатдегідрогенази; цей фермент дає стереоспецифічну реакцію, оскільки завжди утворюється ненасичений (це алкен) транс, тобто фумарат (тоді як цис -ізомер - малеат). Сукцинатдегідрогеназа міститься на внутрішній мембрані мітохондрій, тоді як усі інші ферменти циклу Кребса розсіяні по всьому мітохондрію.
Сукцинатдегідрогеназа має FAD як кофактор; він інгібується оксалоацетатом (інгібування зворотного зв'язку), в той час як він має позитивний модулятор (активатор) сукцинат і фумарат. цикл Кребса потребує енергії, тому єдиною можливістю отримати оксалоацетат від пацієнта є те, що концентрація у пацієнта дуже висока: малат - один з метаболітів з найбільшою концентрацією у клітинах. Реакції, яка перетворює малат в оксалоацетат, також сприяє той факт, що концентрація оксалоацетату утримується низькою під дією цитратсинтази. Реакція, каталізована сукцинатдегідрогеназою, є реакцією самозабезпечення, і це єдиний спосіб здійснити перетворення малату в оксалоацетат.
Концентрація мітохондріального малату повинна бути сумісною з концентрацією цитоплазматичного малату: лише тоді, коли концентрація мітохондріального малату настільки висока, що гарантує перетворення малату в оксалоацетат (у циклі Кребса), тоді малат також можна використовувати у інші шляхи (цитоплазматичні): у цитоплазмі малат може перетворюватися в оксалоацетат, з якого аспартат може бути отриманий під дією GOT (це трансаміназа) або глюкози за допомогою глюконеогенезу.
Повертаємось до сьомої стадії циклу Кребса, вона каталізується ферментом фумарасі: вода додається стереоспецифічним способом для отримання L-малату.
На останній стадії циклу Кребса, про яку ми вже говорили, відбувається дія малатдегідрогеназа. Цей фермент використовує молекулу НАД + для своєї каталітичної дії.
Таким чином, ми завершили опис різних стадій циклу Кребса.
Цикл кребса повністю оборотний.
Для збільшення швидкості циклу Кребса можна збільшити концентрацію метаболітів, присутніх у цьому циклі; одна зі стратегій збільшення швидкості циклу Кребса полягає у перетворенні частини пірувату, що надходить у мітохондрії, в оксалоацетат (під дією піруваткарбоксилази), а не перетворенні всього цього в ацетилкоензим А: таким чином збільшується концентрація оксалоацетату, що є метаболітом циклу Кребса і, отже, збільшує швидкість усього циклу.
У циклі кребса три NAD + перетворюються на три NADH і один FAD у FADH2, і, крім того, отримується GTP: шляхом спрямування зменшувальної потужності, отриманої від циклу Krebs, виробляється подальший АТФ; у дихальному ланцюзі відновлююча сила передається від НАДН і FADH2 до кисню: це перенесення відбувається завдяки ряду ферментів, розташованих на мембрані мітохондрій, які у своїй дії призводять до вироблення АТФ.
Процеси дихального ланцюга є ексергонічними процесами, а звільнена енергія використовується для виробництва АТФ; мета клітини - використати ексергонічні процеси для здійснення синтезу АТФ. Для кожної молекули НАДН, що надходить у дихальний ланцюг, отримують 2,5 молекули АТФ, а для кожної ФАДГ2 - 1,5 молекули АТФ; ця різноманітність пояснюється тим, що FADH2 потрапляє в дихальний ланцюг на нижчому рівні, ніж NADH.
Зі зменшенням сили аеробного метаболізму отримують 30-32 АТФ (219-233 ккал / моль) з ефективністю близько 33% (ефективність анаеробного обміну становить близько 2%).