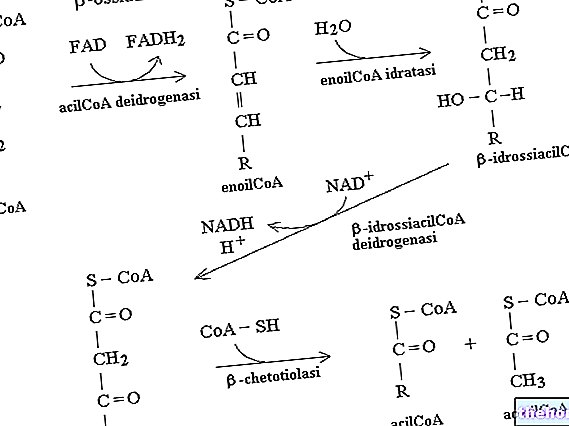
Визначення
Ферменти - це білки, що виробляються в рослинних і тваринних клітинах, які діють як каталізатори прискорення біологічних реакцій без зміни.
Ферменти діють, поєднуючись з певною речовиною, перетворюючи її в іншу речовину; класичні приклади наводяться травними ферментами, присутніми у слині, шлунку, підшлунковій залозі та тонкому кишечнику, які виконують важливу функцію у травленні та допомагають розщеплювати їжу на основні складові, які потім можуть поглинатися та використовуватися організмом, переробляючись іншими ферментами або виводиться у вигляді відходів.
Кожен фермент виконує певну роль: той, який розщеплює жири, наприклад, не діє на білки або вуглеводи. Ферменти необхідні для благополуччя організму. Дефіцит навіть одного ферменту може спричинити серйозні проблеми.Відомим прикладом є фенілкетонурія (ФКУ)-захворювання, що характеризується неможливістю метаболізму незамінної амінокислоти, фенілаланіну, накопичення якої може спричинити фізичні деформації та психічні захворювання.
Біохімічне дослідження
Ферменти - це особливі білки, які мають властивість бути біологічними каталізаторами, тобто вони здатні розщеплювати енергію активації (Eatt) реакції, змінюючи її шлях таким чином, щоб кінетично повільний процес протікав швидше.
Ферменти збільшують кінетику термодинамічно можливих реакцій і, на відміну від каталізаторів, вони більш -менш специфічні: тому вони мають субстратну специфічність.
Фермент не бере участі в стехіометрії реакції: для цього кінцевий каталітичний сайт повинен бути ідентичним вихідному.
У каталітичній дії майже завжди є повільна фаза, яка визначає швидкість процесу.
Коли ми говоримо про ферменти, то неправильно говорити про рівноважні реакції, ми говоримо натомість про стаціонарний стан (стан, при якому певний метаболіт утворюється і споживається безперервно, зберігаючи його концентрацію майже постійною з плином часу). Продукт реакції, каталізованої одним ферментом, зазвичай сам є реагентом для подальшої реакції, каталізується іншим ферментом тощо.
Процеси, що каталізуються ферментами, зазвичай складаються з послідовностей реакцій.
Загальну реакцію, каталізовану ферментом (Е), можна узагальнити таким чином:

Е - фермент
S - підкладка;
ES являє собою аддукт між ферментом і субстратом;
P - добуток;
K - константа швидкості реакції.
Загальний фермент (E) поєднується з субстратом (S), утворюючи аддукт (ES) з постійною швидкістю K1; він може дисоціювати назад на E + S, з постійною швидкістю K2, або, (якщо "живе" досить довго ) можна перейти до форми P з постійною швидкістю K3.
Продукт (Р) може, у свою чергу, рекомбінувати з ферментом і реформувати аддукт з постійною швидкістю К4.
Коли фермент і субстрат змішуються, є частина часу, протягом якої зустріч між двома видами ще не відбулася: тобто існує надзвичайно короткий проміжок часу (який залежить від реакції), протягом якого фермент і субстрат мають ще не виконано; після цього періоду фермент і субстрат у все більшій кількості контактують і утворюється аддукт ES. Згодом фермент діє на субстрат, і продукт вивільняється. Тоді можна сказати, що c " - це початковий часовий інтервал, у якому не можна визначити концентрацію аддукту ES; після цього періоду вважається, що стаціонарний стан встановлено, тобто швидкість процесів, які призводять до отримання аддукту, дорівнює швидкості процесів, які призводять до руйнування аддукту.
Константа Міхаеліса-Ментена (КМ)-це константа рівноваги (посилання на першу рівновагу, описану вище); можна сказати, з хорошим наближенням (оскільки також слід враховувати К3), що КМ представлена співвідношенням між кінетичними константами К2 і К1 (йдеться про руйнування та утворення аддукту ES у першій рівновазі, описаній вище) .
Через константу Михаелиса-Ментена ми маємо "вказівку на спорідненість між ферментом і субстратом: якщо КМ невеликий с"-це "висока спорідненість між ферментом і субстратом, то аддукт ES є стабільним.
Ферменти підлягають регулюванню (або модуляції).
У минулому в основному говорили про негативну модуляцію, тобто інгібування каталітичних можливостей ферменту, але також може бути і позитивна модуляція, тобто існують види, здатні посилювати каталітичні можливості ферменту.
Існує 4 типи гальмування (отримані з наближень, зроблених на моделі для узгодження експериментальних даних з математичними рівняннями):
- гальмування конкуренції
- неконкурентне гальмування
- неконкурентне гальмування
- конкурентне гальмування
Ми говоримо про конкурентне гальмування, коли молекула (інгібітор) здатна конкурувати з субстратом. Для структурної подібності інгібітор може реагувати замість субстрату; звідси і термінологія "конкурентне інгібування". Імовірність того, що фермент зв'язується з інгібітором або субстратом, залежить від концентрації обох речовин та їх спорідненості з ферментом; тому швидкість реакції залежить від цих факторів.
Щоб отримати таку ж швидкість реакції, як і без присутності інгібітора, необхідно мати вищу концентрацію субстрату.
Експериментально показано, що в присутності інгібітора константа Міхаеліса-Ментена зростає.
Що стосується, натомість, "неконкурентного гальмування", взаємодії між молекулою, яка має функціонувати як модулятор (позитивний чи негативний інгібітор), і "ферментом", відбувається у місці, відмінному від того, в якому взаємодія відбувається між ферментом і субстратом; тому ми говоримо про алостеричну модуляцію (від грец аллостерос → інший сайт).
Якщо інгібітор зв'язується з ферментом, він може викликати зміну структури ферменту і, отже, може зменшити ефективність, з якою субстрат зв'язується з ферментом.
У цьому типі процесу константа Міхаеліса-Ментена залишається постійною, оскільки це значення залежить від рівноваги між ферментом і субстратом, і навіть за наявності інгібітора ці рівноваги не змінюються.
Феномен неконкурентного гальмування зустрічається рідко; типовий неконкурентоспроможний інгібітор - це речовина, яка оборотно зв'язується з аддуктом ES, викликаючи ESI:

Інгібування надлишку субстрату іноді може бути неконкурентоспроможним, оскільки це відбувається, коли друга молекула субстрату зв'язується з комплексом ES, що призводить до виникнення комплексу ESS.
З іншого боку, конкурентний інгібітор може зв'язуватися лише з субстратним ферментним аддуктом, як у попередньому випадку: зв'язування субстрату з вільним ферментом викликає конформаційну модифікацію, що робить сайт доступним для інгібітора.
Константа Міхаеліса Ментена зменшується зі збільшенням концентрації інгібітора: очевидно, отже, спорідненість ферменту до субстрату зростає.
Серинова протеаза
Це сімейство ферментів, до яких належать хімотрипсин і трипсин.
Хімотрипсин - протеолітичний та гідролітичний фермент, що розрізає праворуч гідрофобні та ароматичні амінокислоти.
Продукт гена, що кодує хімотрипсин, не активний (він активується за допомогою команди); неактивна форма хімотрипсину представлена поліпептидним ланцюгом з 245 амінокислот. Хімотрипсин має кулясту форму через п’ять дисульфідних містків та інших незначних взаємодій (електростатичні, сили Ван -дер -Ваальса, водневі зв’язки тощо).
Хімотрипсин виробляється хімозними клітинами підшлункової залози, де він міститься в спеціальних мембранах і виводиться через протоку підшлункової залози в кишечник під час перетравлення їжі: хімотрипсин - насправді травний фермент. Білки та поживні речовини, які ми вживаємо з їжею, піддаються перетравленню, щоб зменшити їх до менших ланцюгів, поглинати та перетворювати в енергію (наприклад, амілази та протеази розщеплюють поживні речовини до глюкози та амінокислот, які потрапляють до клітин через кровоносні судини) вони досягають ворітної вени і звідти надходять до печінки, де проходять подальше лікування).
Ферменти виробляються в неактивній формі і активуються лише тоді, коли вони потрапляють на "ділянку, де вони повинні працювати"; як тільки їх дія завершується, вони деактивуються. Після вимкнення фермент не може бути активований повторно: щоб мати "подальшу каталітичну дію, його слід замінити" іншою молекулою ферменту. Якби хімітрипсин вироблявся в активній формі вже в підшлунковій залозі, він би атакував останню: панкреатит - це патологія, спричинена травними ферментами, які вже активуються в підшлунковій залозі (а не в необхідних місцях); деякі з них, якщо їх не лікувати вчасно, привести до смерті.
У хімотрипсині та в усіх серинових протеазах каталітична дія обумовлена наявністю спиртового аніону (-СН2О-) у бічному ланцюзі серину.
Серинові протеази прийняли цю назву саме тому, що їх каталітична дія обумовлена серином.
Після того, як весь фермент виконав свою дію, перш ніж знову мати можливість діяти на субстраті, його слід відновити водою; «вивільнення» серину водою є найповільнішою стадією процесу, і саме ця фаза що визначає швидкість каталізу.
Каталітична дія відбувається у дві фази:
- утворення аніону з каталітичними властивостями (аніон -алкоголят) та подальша нуклеофільна атака на карбонільний вуглець (С = О) з розщепленням пептидного зв’язку та утворенням складного ефіру;
- атака води з відновленням каталізатора (здатний знову проявити свою каталітичну дію).
Різні ферменти, що належать до сімейства серинових протеаз, можуть складатися з різних амінокислот, але для всіх них каталітичний сайт представлений спиртовим аніоном бічного ланцюга серину.
Підродина серинових протеаз - це ферменти, що беруть участь у коагуляції (що полягає у перетворенні білка з їх неактивної форми в "іншу активну форму"). Ці ферменти забезпечують максимально ефективну коагуляцію та її обмеження простір і час (коагуляція повинна відбуватися швидко і повинна відбуватися лише поблизу пошкодженої ділянки) .Ферменти, що беруть участь у коагуляції, активуються каскадно (від активації окремого ферменту отримують мільярди ферментів: кожен активований фермент в свою чергу активує багато інших ферментів).
Тромбоз - це патологія, обумовлена порушенням функцій ферментів згортання крові: вона викликана активацією без необхідності (оскільки немає травм) ферментів, що використовуються при згортанні.
Існують модулюючі (регуляторні) ферменти та інгібуючі ферменти для інших ферментів: взаємодіючи з останніми, вони регулюють або пригнічують їх активність; навіть продукт ферменту може бути інгібітором ферменту. Існують також ферменти, які працюють чим більше, тим більший присутній субстрат.
Лізоцим
Луїджі Пастер виявив, чхаючи на чашку Петрі, що в слизі є фермент, здатний вбивати бактерії: лізоцим; з грецької: лісо = якого розміру; zimo = фермент.
Лізоцим здатний руйнувати клітинну стінку бактерій. Бактерії та взагалі одноклітинні організми потребують механічно стійких структур, які обмежують їх форму; всередині бактерій дуже високий осмотичний тиск, тому вони притягують воду. Плазматична мембрана вибухнула б, якби не було клітинної стінки, яка протистояла б надходженню води і обмежувала б обсяг бактерії.
Клітинна стінка складається з полісахаридного ланцюга, в якому чергуються молекули N-ацетил-глюкозаміну (NAG) та молекули N-ацетил-мурамічної кислоти (NAM); зв'язок між НАГ і НАМ розривається гідролізом. Карбоксильна група NAM у клітинній стінці бере участь у пептидному зв’язку з амінокислотою.
Між різними ланцюжками утворюються містки, що складаються з псевдопептидних зв’язків: розгалуження відбувається за рахунок молекули лізину; структура в цілому дуже розгалужена, і це дає їй високу стійкість.
Лізоцим - це антибіотик (вбиває бактерії): він діє, створюючи тріщину в стінці бактерій; коли ця структура (яка є механічно стійкою) руйнується, бактерія набирає воду, поки вона не лопне. Лізоциму вдається розірвати β-1,4 глюкозидний зв'язок між NAM і NAG.
Каталітичний сайт лізоциму представлений жолобком, що проходить вздовж ферменту, в який вставлений полісахаридний ланцюг: шість глюкозидних кілець ланцюга розміщені в борозенці.
У положенні три канавки с "є дросель: у цьому положенні можна розмістити лише один НАГ, оскільки НАМ, який має вищі розміри, не може потрапити. Фактичний каталітичний майданчик знаходиться між положеннями чотири та п’яте: оскільки існує NAG у позиції три, розріз відбуватиметься між NAM і NAG (а не навпаки); тому вирізання є специфічним.
Оптимальний рН для дії лізоциму - п’ять. У каталітичному центрі ферменту, тобто між положеннями четвертим і п'ятим, є бічні ланцюги аспарагінової кислоти та глутамінової кислоти.
Ступінь гомології: вимірює спорідненість (тобто подібність) між білковими структурами.
Між лізоцимом та лактозосинтазою існує сильний зв’язок.
Лактозосинтетаза синтезує лактозу (яка є основним молочним цукром): лактоза-це галактозил глюкозид, у якому c "є β-1,4-глюкозидним зв'язком між галактозою та глюкозою.
Тому лактозосинтетаза каталізує протилежну реакцію до реакції, що каталізується лізоцимом (який замість цього розщеплює β-1,4 глюкозидний зв’язок)
Лактозосинтетаза є димером, тобто складається з двох білкових ланцюгів, одна з яких має каталітичні властивості та порівнянна з лізоцимом, а інша - регуляторною субодиницею.
Під час вагітності глікопротеїни синтезуються клітинами молочної залози під дією галатозил-трансферази (вона має «гомологію послідовності 40% з лізоцимом): цей фермент здатний переносити галактозильну групу з високоенергетичної структури до структура глікопротеїну. Під час вагітності індукується експресія гена, що кодує галактозил-трансферазу (також існує експресія інших генів, які також дають інші продукти): відбувається збільшення розміру грудей, оскільки він активується молочна залоза (раніше неактивна), яка повинна виробляти молоко. Під час пологів виробляється α-лактальбумін, який є регуляторним білком: він здатний регулювати каталітичну здатність галактозилтрансферази (шляхом розрізнення субстрату). Галактозил-трансфераза, модифікована α-лакталальбуміном, здатна переносити галактозил на молекулу глюкози: утворюючи β-1,4 глікозидний зв’язок і даючи лактозу (лактозосинтетазу).
Отже, трансфераза галактози готує молочну залозу до пологів і виробляє молоко після пологів.
Для отримання глікопротеїнів галактозилтрансфераза зв’язується з галактозилом та NAG; під час пологів лактальний альбумін зв'язується з галактозилтрансферазою, змушуючи останні розпізнавати глюкозу і більше не перетворюючи НАГ, щоб давати лактозу.