"перша частина
Фізіологія
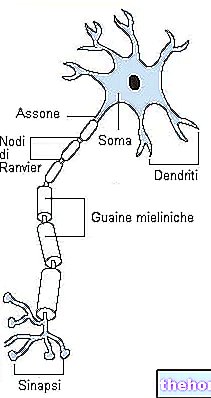
Сенсорні нейрони нервово -м’язового веретена чутливі до розтягування.
Вже в умовах спокою їх проміжна частина достатньо розтягується, щоб спонукати їх посилати нервові імпульси, які вони довіряють чутливим волокнам. У спинному мозку ці волокна скорочуються синапсами безпосередньо з альфа -моторними нейронами, що відповідають за іннервацію тієї ж м’язи, з якої вони почали. Завдяки цій базальній діяльності м’яз, що знаходиться у стані спокою, завжди підтримує певну ступінь напруги, що визначається як «м’язовий тонус».

Під час рухів веретено подовжується і скорочується разом з м’язом. Отже, будь -який жест, що призводить до подовження м’язових волокон, так само впливатиме на веретена, викликаючи збільшення частоти вихідних імпульсів. Ці сигнали негайно переробляються в спинному мозку, викликаючи рефлекторне скорочення м’яза і захищаючи його від пошкоджень, спричинених його надмірним розтягуванням. Ступінь цього рефлекторного скорочення м’язів тим інтенсивніше, чим більша частота нервових імпульсів (у свою чергу, прямо пропорційна ступеню розтягування, сприйнятому чутливими волокнами нервово -м’язового веретена).
Одночасно з активацією альфа -моторних нейронів інгібуючі сенсорні волокна активують гальмівні інтернейрони, відповідальні за тимчасове «заглушення» альфа -моторних нейронів, які іннервують м’язи -антагоністи, запобігаючи їх скорочення.
Все це відбувається за допомогою мимовільного механізму, званого рефлексом розтягування або міотатичним рефлексом [да міо = м'яз e tasis = розтягування].
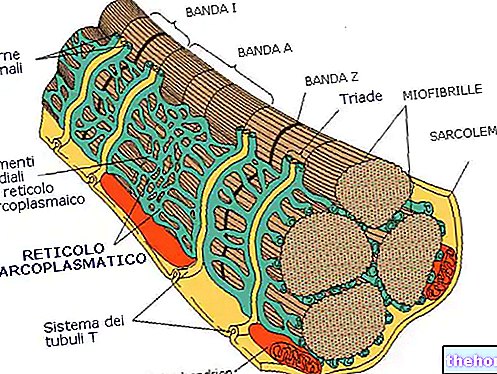
Роль гамма -моторних нейронів зараз залишається пояснити. Їх завдання - регулювати чутливість нервово -м’язових веретенів відповідно до ступеня розтягування, щоб вони залишалися активними навіть при скороченні м’язів. Все це стає можливим завдяки так званій альфа-гамма-коактивації, тобто одночасному скороченню фузальних та інтрафузальних волокон. Оскільки останні іннервуються на обох кінцях м’язів, їх скорочення призводить до подовження центральної області, яка утримує сенсорні закінчення розтягнутими.
У той час як м’яз, іннервований альфа-моторним нейроном, скорочується за рахунок зменшення натягу на веретеноподібну капсулу, одночасна активація інтрафузальних волокон, гарантована гама-коактивацією, утримує нервово-м’язовий веретено активним. Таким чином, чутливість рецептора може залишатися постійною на кожному рівні скорочення, забезпечуючи більшу плавність рухів та швидшу реакцію м’язів у разі потреби.
Подальше дослідження активності нервово -м’язових веретенів накладає класифікацію волокон з мішків ядер на два підкласи: статичних волокон мішечка та волокон динамічних мішечків. Останні, іннервовані сенсорними волокнами типу I, отримують переважно різкі та швидкі зміни швидкості , також завдяки «руховій іннервації, що складається з особливо швидких гамма -волокон (багатомієлінованих, званих динамічними гамма -волокнами).
Хоча щойно описані волокна ідеально підходять для швидкого зчитування змін довжини м’язів, статичні волокна мішечка надають більш точну інформацію про тривалість та ступінь зміни напруги (вони іннервуються статичними волокнами гамма -II). Розряд вторинних волокон отримує ЦНС інформація про "ступінь" подовження м'язів і, через первинні, про швидкість подовження.
Нарешті, слід зазначити, що на активність нервово -м’язових веретенів впливають численні позазастосовні фактори, такі як температура (тепло знижує їх активність, викликаючи розслаблення, а холод збільшує жорсткість) та ступінь втоми (ефективність міотатичного рефлексу l » зменшується, коли спортсмен втомився, схильний до ризику травми м’язів).