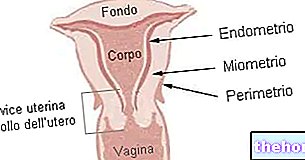
Для того, щоб мати можливість зрозуміло говорити про "гемоглобін (Hb), корисно подбати про перше міоглобін (Мб) який дуже схожий на гемоглобін, але набагато простіший. Між гемоглобіном і міоглобіном існують близькі споріднені стосунки: обидва є кон'югованими білками, а їх протетична група (небілкова частина)-це група гема.
Міоглобін - це глобулярний білок, що складається з єдиного ланцюга з приблизно ста п’ятдесяти амінокислот (він залежить від організму) і його молекулярна маса становить близько 18 Кд.
Як згадувалося, він обладнаний гемовою групою, яка вставлена в гідрофобну (або ліпофільну) частину білка, що складається зі складок, що відносяться до структур α-спіралі волокнистих білків.
Міоглобін в основному складається з сегментів α-спіралей, присутніх у кількості восьми і майже виключно складається з неполярних залишків (лейцин, валін, метіонін та фенілаланін), тоді як полярні залишки практично відсутні (аспарагінова кислота, глутамінова кислота, лізин) та аргінін); єдиними полярними залишками є два гістидини, які відіграють фундаментальну роль у приєднанні кисню до групи гему.
Група гему є хромофорною групою (поглинається у видимому) і є функціональною групою міоглобіну.
Дивіться також: глікований гемоглобін - гемоглобін у сечі
Трохи хімії

Зв'язок між протопорфірином і залізом є типовим зв'язком координаційних сполук, які є хімічними сполуками, в яких центральний атом (або іон) утворює зв'язки з іншими хімічними речовинами в кількості, більшій за його число окислення (електричний заряд). У разі гему ці зв’язки є оборотними і слабкими.
Координаційне число (кількість координаційних зв’язків) заліза дорівнює шість: навколо заліза може бути шість молекул, які поділяють зв’язуючі електрони.
Щоб утворити координаційну сполуку, потрібні дві орбіталі з правильною орієнтацією: одна здатна "придбати" електрони, а інша здатна віддати їх.
У гемі залізо утворює чотири площинні зв’язки з чотирма атомами азоту в центрі прото-порфіринового кільця та п’ятий зв’язок із проксимальним азотом гістидину; залізо має шостий вільний координаційний зв’язок і може зв’язуватися з киснем.
Коли залізо має форму вільного іона, його тип є орбіталями d всі вони мають однакову енергію; у міоглобіні іон заліза зв’язаний з протопорфірином та гістидином: ці види магнітно збурюють орбіталі d трохи заліза; ступінь збурення буде різною для різних орбіталей d в залежності від їх просторової орієнтації та виду збудливих видів. Оскільки загальна енергія орбіталей повинна бути постійною, збурення спричиняє енергетичний поділ між різними орбіталями: енергія, яку отримують одні орбіталі, еквівалентна енергії, втраченої іншими.
Якщо поділ між орбіталями не дуже великий, краще електронне розташування з високими спінами: зв'язуючі електрони намагаються розташуватися в паралельних спінах на якомога більшій кількості рівнів (максимальна кратність); якщо, навпаки, збурення дуже сильне і між орбіталями існує велике роз’єднання, може бути зручніше об’єднати електрони зв’язку на нижчих енергетичних (низькоспінових) орбіталях.
Коли залізо зв'язується з киснем, молекула приймає низьку спінову структуру, тоді як у заліза вільний шостий координаційний зв'язок, молекула має високу спінову структуру.
Завдяки цій спіновій різниці за допомогою спектрального аналізу міоглобіну ми можемо зрозуміти, чи зв'язаний з ним кисень (MbO2) чи ні (Mb).
Міоглобін - типовий м’язовий білок (але він не міститься тільки в м’язах).
Міоглобін витягується з кашалота, в якому він присутній у великій кількості, а потім очищається.
Китоподібні мають дихання, подібне до людського: маючи легені, вони повинні поглинати повітря через дихальний процес; кашалот повинен доставляти якомога більше кисню в м’язи, які здатні накопичувати кисень, зв’язуючи його з міоглобіном, що знаходиться в них; потім кисень повільно вивільняється, коли китоподібний занурюється, оскільки для його метаболізму потрібен кисень: тим більша кількість кисню, який кашалот здатний поглинути, і тим більше кисню доступно під час занурення.
Міоглібін пов'язує кисень оборотним способом і присутній у периферійних тканинах у більшому відсотку, чим більше ця тканина звикла працювати з віддаленими у часі запасами кисню.
<--- Міоглобін-це білок, присутній у м’язах, функція якого якраз і є функцією «резервуару» кисню.

Те, що робить м’ясо більш -менш червоним, - це вміст гемопротеїнів (саме гем робить м’ясо червоним).
Гемоглобін має багато структурних подібностей з міоглобіном і здатний зв’язувати молекулярний кисень оборотним шляхом; але, хоча міоглобін обмежений м’язами та периферичними тканинами в цілому, гемоглобін міститься в еритроцитах або еритроцитах (це псевдоклітини, тобто вони не є справжніми клітинами), які складають 40% крові.
На відміну від міоглобіну, робота гемоглобіну полягає в тому, щоб взяти кисень у легенях, вивільнити його в клітини, де він необхідний, взяти вуглекислий газ і випустити його в легені, де цикл починається знову.
L "гемоглобін це тетраметр, тобто складається з чотирьох поліпептидних ланцюжків, кожен з яких має гемову групу і однакових по два (у людини є два альфа -ланцюги та два бета -ланцюга).
Основною функцією гемоглобіну є транспортування кисню; інша функція крові, в якій бере участь гемоглобін, - транспортування речовин до тканин.
На шляху від легенів (багатих киснем) до тканин гемоглобін переносить кисень (одночасно інші речовини потрапляють до тканин), тоді як на зворотному шляху він переносить відходи, зібрані тканинами, особливо вуглець діоксид, що утворюється при метаболізмі.
У розвитку людини є гени, які експресуються лише протягом певного періоду часу; з цієї причини існують різні гемоглобіни: фетальний, ембріональний, дорослого чоловіка.
Ланцюги, що складають ці різні гемоглобіни, мають різну структуру, але з деякою подібністю насправді їхня функція більш -менш однакова.
Пояснення наявності кількох різних ланцюжків таке: в ході еволюційного процесу організмів навіть гемоглобін еволюціонував, спеціалізуючись на транспортуванні кисню з областей, багатих ним, до областей з дефіцитом. На початку еволюційного ланцюга "гемоглобін транспортував кисень у невеликих організмах; в ході еволюції організми досягали більших розмірів, тому гемоглобін був змінений, щоб мати можливість транспортувати кисень до областей, що знаходяться подалі від точки, де він багатий; для цього вони були закодовані в ході еволюційного процесу новими структурами ланцюгів, що складають гемоглобін.
Міоглобін зв'язує кисень навіть при помірному тиску; у периферичних тканинах існує тиск (PO2) близько 30 мм рт.ст.: міоглобін при цьому тиску не виділяє кисень, тому він буде неефективним як носій кисню. З іншого боку, гемоглобін , він має більш еластичну поведінку: він зв'язує кисень з високим тиском і виділяє його, коли тиск зменшується.
Коли білок функціонально активний, він може дещо змінити свою форму; наприклад, кисневий міоглобін має іншу форму, ніж неоксигенований міоглобін, і ця мутація не впливає на сусідів.
Інша ситуація у випадку з асоційованими білками, такими як гемоглобін: коли ланцюг насичується киснем, він викликає зміну своєї форми, але ця модифікація є тривимірною, тому інші ланцюги тетраметра також зазнають впливу. один з одним., припускає, що модифікація одного впливає на інших сусідів, навіть якщо в різній мірі; коли ланцюг насичується киснем, інші ланцюги тетраметра приймають "менш вороже ставлення" до кисню: складність, з якою це ланцюг оксигенати зменшуються, оскільки ланцюги, близькі до нього, по черзі окислюються. Те ж саме стосується дезоксигенації.
Четвертинна структура дезоксигемоглобіну називається формою Т (напружена), тоді як структура оксигемоглобіну називається формою R (вивільняється); у напруженому стані існує низка досить сильних електростатичних взаємодій між кислотними амінокислотами та основними амінокислотами, які призводять до жорсткої структури дезоксигемоглобіну (ось чому "напружена форма"), тоді як коли кисень пов'язаний, сутність цих взаємодії зменшуються (звідси "випущена форма"). Крім того, за відсутності кисню заряд гістидину (див. Структуру) стабілізується протилежним зарядом аспарагінової кислоти, тоді як у присутності кисню білок втрачає протон; все це передбачає, що кисневий гемоглобін є сильнішою кислотою, ніж дезоксигенований гемоглобін: ефект Бора.
Залежно від рН, група гему більш -менш легко зв’язується з киснем: у кислому середовищі гемоглобін легше виділяє кисень (напружена форма стабільна), тоді як у базовому середовищі зв’язок з киснем міцніший.

Кожен гемоглобін виділяє 0,7 протонів на моль кисню (O2), що надходить.
Ефект Бора дозволяє гемоглобіну покращити свою здатність переносити кисень.
Гемоглобін, який рухається від легенів до тканин, повинен врівноважуватися як функція тиску, рН та температури.
Давайте подивимося вплив температури.
Температура в легеневих альвеолах приблизно на 1-1,5 ° C нижче зовнішньої температури, тоді як у м’язах температура становить приблизно 36,5-37 ° C; із збільшенням температури коефіцієнт насичення падає (при тому ж тиску): це відбувається тому, що кінетична енергія збільшується і сприяє дисоціації.

Існують і інші фактори, які можуть впливати на здатність гемоглобіну зв'язуватися з киснем, одним з яких є концентрація 2,3 бісфосфогліцерату.
2,3 бісфосфогліцерат є метаболічним речовиною, присутнім в еритроцитах у концентрації 4-5 мМ (ні в одній іншій частині організму він не присутній у такій високій концентрації).
При фізіологічному рН 2,3 бісфосфогліцерат депротонується і на ньому є п’ять негативних зарядів; він вклинюється між двома бета -ланцюгами гемоглобіну, оскільки ці ланцюги мають високу концентрацію позитивних зарядів. Електростатичні взаємодії між бета -ланцюгами та 2,3 -бісфосфогліцератом надають системі певну жорсткість: виходить напружена структура, яка має невелику спорідненість до кисню; під час оксигенації 2,3 -бісфосфогліцерат потім видаляється.
В еритроцитах c "-це спеціальний апарат, який перетворює 1,3 бісфосфогліцерат (вироблений шляхом метаболізму) у 2,3 бісфосфогліцерат так, що він досягає концентрації 4-5 мМ, а отже, гемоглобін здатний обмінювати" кисень у тканинах.
Гемоглобін, що надходить до тканини, знаходиться у вивільненому стані (зв’язаному з киснем), але поблизу тканини він карбоксилюється і переходить у напружений стан: білок у цьому стані має меншу схильність зв’язуватися з киснем до стану, що вивільнився, тому гемоглобін виділяє кисень до тканини; крім того, в результаті реакції між водою та вуглекислим газом відбувається виробництво іонів Н +, отже, додатковий кисень через ефект Бора.
Вуглекислий газ дифундує в еритроцит, що проходить через плазматичну мембрану; оскільки еритроцити складають близько 40% крові, слід очікувати, що лише 40% вуглекислого газу, що дифундує з тканин, потрапляє до них, насправді 90% вуглекислого газу надходить до еритроцитів, оскільки вони містять фермент, який перетворює вуглекислий газ в вугільній кислоті це призводить до того, що нерухома концентрація вуглекислого газу в еритроцитах низька, а отже, швидкість надходження висока.

Ще одне явище, яке відбувається, коли еритроцит досягає тканини, таке: за градієнтом "НСО3- (похідне вуглекислого газу) виходить з" еритроциту і, щоб збалансувати вихід негативного заряду, ми маємо "надходження хлоридів, які визначає збільшення осмотичного тиску: щоб збалансувати цю зміну, також відбувається надходження води, що викликає набряк еритроцита (ефект ГАМБУРГЕРА.). Тому венозні еритроцити (спрямовані до легенів) більш круглі, ніж артеріальні.