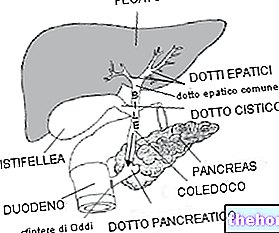
Дивіться також: кетогенна дієта; діабетичний кетоацидоз.
Загальність
Раніше вважалося, що кетонові тіла були викликані надмірним обміном речовин, викликаним надмірним вживанням жиру або цукровим діабетом. Кетонові тіла, з іншого боку, природним чином виробляються нашим тілом: мозок адаптується до використання цих метаболітів в умовах тривале голодування (у хворих на цукровий діабет кетонові тіла замінюють метаболізм глюкози) Крім того, може статися роздратування шляху кетонових тіл у разі неправильного харчування.
Що таке кетонові тіла
Кетонові тіла є похідними ліпідів (вони походять від метаболізму ліпідів, майже виключно печінкових), але мають властивості, що роблять їх схожими на цукор:
- Висока швидкість введення;
- Швидкий у використанні.
Навіть деякі амінокислоти, зокрема метаболічні умови, можуть походити від кетонових тіл (лейцин, лізин, фенілаланін, ізолейцин, триптофан та тирозин).
Біологічна роль
- Кетонові тіла мають невеликі розміри, тому транспортуються дуже швидко (набагато більше, ніж жирні кислоти, які, з іншого боку, потребують транспортних білків, таких як альбумін);
- кетонові тіла використовуються майже виключно м’язами та периферійними тканинами, а також серцем (20-30% енергії, яку він використовує, надходить з кетонових тіл) та мозком (у разі тривалого голодування).
Синтез
Кетонові тіла синтезуються ацетил -коферментом А, який походить від метаболізму жирних кислот.
Фермент, який каталізує першу стадію, - це Β-кетотіолаза, який використовує сірку ацетил-коферменту А для утворення Β-кетоацил-коферменту А (це протилежна реакція, що спостерігається при Β-окисленні жирних кислот); ця реакція не є спонтанною, а обумовлена наступною реакцією , каталізований з "гідроксиметилглутаріл кофермент А синтаза і який передбачає приєднання другого ацетил-коферменту А, одержуючи 3-гідрокси-3-метилглутарил-кофермент А.
Згодом втручається літичний фермент, який перетворює 3-гідрокси 3-метилглутаріл-кофермент А в оцтовий оцет, який є кетоновим тілом. Ацетат оцту може бути направлений до периферійних тканин або під дією ферменту гідроксибутиратдегідрогеназа, перетворюється на 3-Β-гідроксибутират. Якщо ацетат оцту в дуже високій концентрації, він також може спонтанно декарбоксилюватися до ацетону.
Ацетон, ацетат оцту та 3-Β-гідроксибутират-це три кетонові тіла, які ми розглядаємо; ацетон - це відхідний продукт, який утворюється випадковим чином на шляху кетонових тіл і видаляється шляхом видиху та дихання.
Застосування в периферичних тканинах
Кетонові тіла, що виробляються в печінці, надходять до периферичних тканин.
Тепер подивимося, що станеться, коли ацетат оцту та 3-Β-гідроксибутират потрапляють до периферійних тканин. ацетилкоензим А: отже, необхідно перетворити Β-кетокислоту на Β-кетоациловий кофермент А.
Коли ацетат оцту надходить у мітохондрію клітини периферійної тканини, він піддається дії ферменту сукциніл -коферменту А трансферази: за допомогою цього ферменту ацетат оцту реагує з сукциніловим коферментом А (що надходить із циклу Кребса) та отримують сукцинат та оцтовий ацетилкофермент А.
Використовуючи сукциніловий кофермент А, щоб активувати ацетат оцту, ми переходимо до циклу Кребса, стадії, яка виробляє ГТФ: це процес, з точки зору енергії, який клітина готова заплатити, щоб отримати коензим А ацетилового оцту ; останній потім переходить під дію Β-кетотіолаза (Фермент Β-окислення) для утворення двох молекул ацетил-коферменту А, які надходять у цикл Кребса.
Якщо 3-Β-гідроксибутират надходить до периферійних тканин, останній всередині мітохондрії перетворюється в оцтовий ацетон під дією ферменту Β-гідроксибутиратдегідрогенази з виробленням НАДН, що відповідає приблизно 2,5 АТФ; одержаний ацетат оцту слідує описаному вище шляху.
Клітина периферійної тканини отримує більше енергії з 3-Β-гідроксибутирату, а не з ацетату оцту, але доставка того чи іншого в периферичні тканини залежить від енергетичної доступності печінки.
C "-це незначна кількість метаболізованих жирних кислот, що містяться в пероксисомах, а не в мітохондріях; пероксисоми-це органели, менші за мітохондрії та багаті іонами металів та ферментами пероксидази. Ферменти пероксидази використовують перекис водню для сприяння окисно-відновним процесам, отже, у пероксисомах є являє собою ферментативну систему, здатну виробляти перекис водню.
При Β-окисленні в пероксисомах "ацильний кофермент А отримують дією"ацил -кофермент А оксидаза (З іншого боку, у мітохондріях діяв фермент ацил -кофермент А -дегідрогеназа.) Також у цьому випадку утворюється транс 2,3 -еноїльний кофермент А, який зазнає дії біфункціонального ферменту (він виконує ту ж функцію, що і в мітохондріях за допомогою "еноїл-коферменту А-гідратази та L-Β-гідроксиацил-коферменту А-дегідрогенази) і таким чином перетворюється на Β-кетоацил-кофермент А. Цей останній, як і в мітохондріях, зазнає дії Β-кетотіолази та ацетилкоензиму А а ацильний кофермент А отримують з вуглецевим скелетом, зменшеним на дві одиниці порівняно з вихідним, який повертається в циркуляцію.